


The dynamic N1-methyladenosine methylome in eukaryotic messenger RNA
近些年研究表明N6-methyladenosine (m6A) 在mRNA代谢中扮演重要角色,此外,pseudouridine (Ψ)和 5-methylcytosine 也被发现在基因表达的转录后修饰中启着重要作用。N1-methyladenosine在信使RNA中的修饰则未见报道。
We translate sequences to science and industry.
亚硫酸测序的优点使其能够应用于基因组尺度的分析,然而在此之前,总体测量DNA甲基化仅能通过其他技术手段实现,如限制性标记基因组扫描(Urich et al. 2015)。在人类基因组计划完成后,许多科学家随后绘制出了人类表观组图谱,这一表观遗传信息的获得将对遗传序列执行和调控功能的理解十分重要,表观组比基因组相对不稳定,因此表观组被认为在基因-环境互作中起重要作用(Schübeler 2015; Law & Holland 2019)。

.png)

近些年研究表明N6-methyladenosine (m6A) 在mRNA代谢中扮演重要角色,此外,pseudouridine (Ψ)和 5-methylcytosine 也被发现在基因表达的转录后修饰中启着重要作用。N1-methyladenosine在信使RNA中的修饰则未见报道。
1. DNA 甲基化是一种重要的表观遗传调控机制,在众多物种中被广泛研究。 2.以前的研究认为在哺乳动物中只存在5mC 甲基化,N6-mA甲基化只在原核生物和少数低等真核生物存在。 3.哺乳动物是否存在N6-mA甲基化,一直缺乏直接而有力的证据。
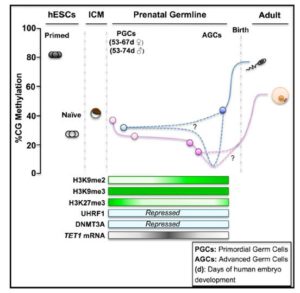
精子和卵子的融合是动物胚胎发育的起点,不过这一过程并不仅仅是简单的融合。哺乳动物细胞的甲基化模式是在发育中逐渐形成的,在胚胎发育初期,全基因组的甲基化模式会发生明显变化。比如说小鼠胚胎受精之前,雌雄生殖细胞具有较高的甲基化水平,胚胎着床前发生大规模的DNA去甲基化。

长链非编码RNAs(lncRNA)是一类长度大于200nt,不编码蛋白质的RNA分子。与mRNAs相识,lncRNA也是由RNA聚合酶II转录,经历剪接和多聚腺苷酸化。根据它们与蛋白质编码基因的相对定位,lncRNA可分为反义转录物(antisense transcripts)、长链基因内非编码RNAs(long intronic noncoding RNAs)和长链基因间非编码RNAs(long intergenic noncoding RNAs, lincRNAs)。有一些lincRNA已被证明在多种生物学过程如剂量补偿、转录调控、表观遗传调控和细胞多能性维持等中发挥重要作用。以往的研究也证实lincRNA在脂肪生成和肌肉组织发育中发挥作用。

表观遗传调控是细胞核内染色体的高级组织形式,是目前干细胞研究领域的前沿和热点问题。包括胚胎干细胞和诱导干细胞在内的多功能干细胞都可以利用遗传学和表观遗传学的这种复杂网络,来维持自我更新和多向分化之间的精密平衡